
La
phospholipase A2 coupe la liaison ester en position 2 des phospholipides
(1,2-diacylphosphoglycérides), libérant un acide gras et un lysophospholipide.
Examen de l'élément de module 'Relation sructure-fonction dans le métabolisme primaire et éléments de base du métabolisme secondaire des plantes', module 'Biochimie, Génétique et Amélioration des Plantes' du Parcours S5 : 'Biologie Appliquée aux Productions Végétales', Novembre 2012, Durée : 1 heure 15 minutes .......... Corrections
brèves
QUESTION 1
Les phospholipases A1, A2, C et D sont des enzymes qui hydrolysent les phospholipides.

A. La phospholipase A2 qui a été découverte, d'abord dans les venins de serpents et d'abeilles puis dans les tissus de mammifères et des plantes (protection contre les pathogènes), coupe la liaison ester en position 2 des 1,2-diacylphosphoglycérides, libérant un acide gras et un lysophospholipide. La réaction catalysée par la phospholipase A2 cytosolique implique la présence d'ions calcium (Ca++). Pour étudier le mécanisme cinétique de cette réaction, les vitesses initiales d'hydrolyse de la dibutyryl-lécithine (DBL) ont été mesurées à différentes concentrations de ce dernier et d'ion Ca++ (substrats, S). L'activité enzymatique est déterminée en effectuant le titrage par la soude de l'acide libéré. Les vitesses sont exprimées en µmoles d'acide libérées par min et par mg de protéine. Les cinétiques 1/v = f(1/(S)) (représentations de Lineweaver-Burk) sont résumées dans les figures 1 et 2 ci-dessous. Les diagrammes secondaires tracés à partir des diagrammes primaires, représentent les variations des intersections des droites 1/v = f(1/(S)) avec l'axe des ordonnées (b) en fonction des inverses des concentrations du substrat maintenu constant.
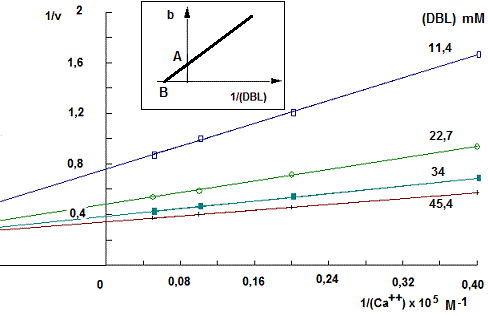
Fig. 1

Fig. 2
Les
droites de la représentation 1/v = f(1/(Ca++)) font intersection au dessus de l'axe des abscisses en un point dont la projection sur cet axe est - 0,25 x 105 M-1. Les points d'intersections avec les différents axes des diagrammes secondaires sont respectivement
A = 0,2 µmole-1 x min x mg (= C) et B = - 0,031 mM-1.
1.
En se basant uniquement sur la cinétique, proposer un ou plusieurs
mécanisme(s) de catalyse susceptible(s) de rendre compte de
toutes les cinétiques de la phospholipase A2 obtenues avec
la dibutyryl-lécithine (DBL) et le calcium.
2. Donner le(s) schéma(s) de Cleland général(aux)
correspondant(s).
B. Afin de confirmer le mécanisme de catalyse de la phospholipase
A2, les effets inhibiteurs de l'acide butyrique et du baryum ont été
étudiés. Ils ont permis d'obtenir les cinétiques résumées dans la figure 3.

Fig. 3
1.
Préciser les types d'inhibition exercées par le baryum vis-à-vis du calcium et de la DBL.
2. De quels substrat baryum est-il analogue ?. Justifier.
3. Préciser les types d'inhibitions exercées par l'acide butyrique vis-à-vis du calcium et de la DBL.
4. De quel substrat l'acide butyrique est-il analogue ? Justifier.
5. En s'aidant des types d'inhibition obtenus avec les analogues
des substrats, déterminer le mécanisme de catalyse précis
de la phospholipase A2. Justifier la confirmation ou l'infirmation
de tout mécanisme proposé. Calculer les paramètres
cinétiques de la phospholipase A2.
6. Proposer un schéma de Cleland pour le mécanisme
de catalyse de la phospholipase A2.
QUESTION 2.
Interpréter brièvement les structures enzymatiques, relatives à la phospholipase A2 et illustrées par les figures 4 et 5 suivantes.
Fig. 4
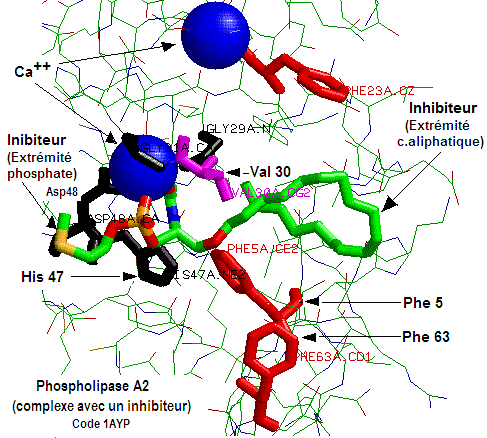
Fig. 5
Corrections
brèves
Question 1:
A.1. Interprétation des cinétiques de la phospholipase A2
- Avec Ca++ : Les diagrammes primaires sont sous formes de faisceaux de droites faisant
intersection à gauche de l'axe des ordonnée et au dessus de l'axe des abscisses:
- Avec la dibutyryl-lécithine (DBL), la cinétique
1v = f(1/(DBL) est sous forme de droites faisant intersection sur l'axe des ordonnée.
Cescinétiques sont en faveur des mécanismes de catalyse:
1/ Mécanisme
au hasard fixation dépendante et 2/ Mécanisme
Séquencé.
A.2.Schéma(s) de Cleland général(aux) correspondant(s)
aux deux mécanisme: voir le cours
B.1. Préciser les types d'inhibition exercées par le
baryum vis-à-vis du calcium et de la DBL: Baryum: inhibiteur
compétitif vis à vis du calcium et de la DBL.
B.2. Baryum est un analogue structural du calcium. Même s'il compétitif vis à vis de la DBL, il ne peut être analogue structural de celle ci (structure différente)
B.3. Acide butyrique: inhibiteur
incompétitif vis à vis du Ca++ et compétitif vis à vis de la DBL
B.4. L'acide butyrique est analogue structural de de la DBL.



Réponse à la Question 1 (suite):
B.5. En s'aidant des types d'inhibition obtenus avec les analogues des substrats, le mécanisme de catalyse précis de la phospholipase A2 est un mécanisme séquencé (ordonné) où le substrat directeur est le Ca++. En effet, l'inhibition incompétitive exercée par l'acide butyrique (analogue de la DBL) vis à vis du Ca++ est expliquée par l'absence de fixation de la DBL sur l'enzyme libre. Cette fixation est conditionnée par la fixation du calcium. Le mécanisme au hasard fixation dépendante ne peut être retenu, carl'inhibition par les analogues de substrats est de type mixte. Le mécanisme de catalyse de la phospholipase A2 est un mécanisme séquencé avec les paramètres cinétiques; KsCa = 0,04 mM, KmDBL = 32,26 mM et Vmax = 5 µmole/min/mg.Réponse à la Question 2:
La
figure 4 montre que la phospholipase A2 est une enzyme à Ca++.
L'extrêmité hydrophile du phospholipide est dirigée
vers le Ca++ alors que la partie hydrophobe (chaines carbonnées
d'acides gras) est orientée vers les résidus aromatiques
des acides aminés de l'enzyme (Tyr, Trp, Phe). Cette structure
montre l'importance des interactions hydrophobes dans la fixation
et l'orientation des phospholipides, substrats de la phospholipase
A2.
La figure 5. Le calcium est lié dans le site actif par l'acide
aspartique (Asp48); L'extrêmité phosphate de l'inhibiteur est orientée vers le Ca++, alors que l'extrêmité aliphatique est orientée vers des résidus phénoliques
des phénylalanine (Phe5, Phe63). D'où la mise en jeu
d'interactions hydrophobes.
Pour rappel, la phospholipase A2 coupe les acides gras en position sn-2
des phospholipides. Cette position contient souvent des acides gras
polyinsaturés (plusieurs doubles liaisons comme l'acide
arachidomique) qui, une fois libérés, contribuent
à la formation de plusieurs eicosanoïdes comme les prostaglandines,
agent de l'inflammation. La cyclooxygénase
(COX) est derrière cette action par un processus de peroxydation.
L'aspirine (acide acétylsalicylique), inhibiteur irréversible
de la COX, bloque la formation des prostaglandine et met fin à
l'inflammation.
Liens utiles:
- Enzymologie-enzymes (exercices)
- QCM-Enzymes. Structure et fonction (bases)
- Hexokinase
- Coenzyme
Atransférase
- Amine
oxydase
- Inhibiteurs.
Applications
- QCM-Inhibiteurs-1
- QCM-Inhibiteurs-2.
- Phospholipase, Allostérie
- Citrate synthétase, Peroxydase (Examen)
- Chélatase, Aspartate transcarbamylase. Examen
- Polyphénoloxydase. Examen
- Citrate synthétase. Examen
- Shikimate déshydrogénase. Examen
- Chelatase, GOT, PPO, désaturase. Examen
- Carboxylestérase, PAL. Examen S5, 2013
- Pyrophosphorylase, Méthylestérase. Examen S5, 2013
Ouvrages de la préparation de la transition Secondaire-Supérieur


-
من علوم الحياة إلى البيوكيمياء
Des Sciences de la vie au Lycée à la Biochimie à l'Université
-
تحميل ملفات في مواضيع البيوكيمياء
Téléchargement de fichiers en Biochimie

BIOCHIMIE. TP CONSIGNES (Vidéos)
Chaine Youtube (abonnement). Plusieurs vidéos multilingues

مجلة إخبارية - Newsletter

ANNONCES, INFORMATIONS
Congrès, Conférences,...
- Congrès, Conférences
- Cours, Workshops
- Bourses, Postdocs
- Biotech-ecolo annonces
CONCOURS:
Concours de Pharmacie
Afin de pouvoir continuer à servir les visiteurs, soutenez nos actions sur le site takween en faisant acquisition des ouvrages et supports pédagogiques desitinés à améliorer l'enseignement et la recherche scientfique en Biochimie.


Faculty of Sciences, Cadi Ayyad University
Marrakech, 40000, Morocco
Email: baaziz@uca.ac.ma
Phone: 212524434649 (post 513)
Fax: 212524434669
About takween.com