
| French |
Arabic
|
English
|
| Enzymes | إنزيمات | enzymes |
| Site actif | موقع نشيط | Active site |
Connaître comment les enzymes (catalyseurs biologiques) assurent la catalyse renvoie à l'élucidation de leur mécanisme d'action qui implique le contact du substrat avec l'enzyme à travers un site actif.
Le site actif est constitué de quelques acides aminés. Le suivi de la catalyse homogène permet d'élucider en détail le comportement des enzymes et déterminer les paramètres cinétiques correspondants.
---> Vidéo 'Enzymes et catalyse biologique - الإنزيمات، محفزات بيولوجية
Site actif des enzymes (protéines de catalyse bioligique)
LA CATALYSE ENZYMATIQUE PASSE PAR LA FORMATION d'un SITE ACTIF.
Le site actif des enzymes est une région privilégiée de l'enzyme qui interagit avec les substrats. Le site actif des enzymes est constitué d'acides aminés (AA) de 4 types:
1/ AA 'de contact', 2/ AA 'auxiliaires', 3/ AA 'collaborateurs' et 4/ AA 'non collaborateurs'.

Ce sont les groupements fonctionnelles des radicaux (R) des AA du site actif des enzymes qui interviennent dans la catalyse. Ces groupements sont peu nombreux. Il s'agit le plus souvent des résidus SERYL, ASPARTYL ou GLUTAMYL, CYSTEYL, HISTIDYL LYSYL et accessoirement ARGINYL et TYROSYL (QCM catalyse et site actif des enzymes). La capacité des AA du site actif des enzymes à participer à la catalyse est liée à leur aptitude à transférer des électrons (caractère d'agents nucléophiles) et des protons (caractère acide-base selon Broensted).
Le caractère de nucléophilie est plus prononcé pour la sérine, la cystéine et l'acide aspartique.
Le caractère acide-base est plus prononcé pour l'histidine et la tyrosine.
L'histdine intervient comme relais de proton où le noyau imidazole joue le rôle de 'renforceur' du caractère de nucléophilie des donneurs d'électrons (substrats) en captant le proton de leur OH.
Ce sont finalement que quelques acides aminés de l'enzyme (AA de contact) qui sont nécessaires pour la catalyse. L'élimination de certains acides aminés n'entraîne pas la disparition de l'activité enzymatique.
Exemple du site actif des enzymes. Cas du lysozyme (figure ci-dessous). Le lysozyme (muramidase) est une enzyme de défense contre les infections bactériennes . Elle est constituée de 130 acides aminés chez l'Homme. Elle est présentes dans plusieurs sécrétions (salive, mucus, larmes, lait maternel...) et dans le blanc d'œuf (129 acides aminés chez la poule)
Structures fonctionnelles des enzymes (protéines de catalyse biologique)
La structure tertiaire (ou tridimensionnelle ou conformation) correspondant au repliement de la protéine enzymatique est une structure primordiale nécessaire dans la catalyse. Elle permet la formation du site actif par rapprochement des acides aminés le constituant (ex AA 57, AA195, AA220). Cette conformation joue, également un rôle dans la protection du site actif. La diffraction aux rayons X a montré que l'organisation tridimensionnelle du site actif des enzymes n'est pas tout à fait la même suivant l'absence ou la présence du substrat. Ainsi, était née la théorie dite de 'Conformation induite' (induced fit) où le site actif est perçu comme un 'ensemble souple' en s'adaptant exactement à la conformation du substrat. L'une des preuves expérimentales appuyant cette théorie réside dans les changements de spectres d'absorption et de fluorescence de certaines enzymes en présence ou en absence des substrats.
La structure quaternaire correspond à l'association de l'enzyme en sous unités (QCM-Protéines). Elle donne les derniers ajustements conformationnels. Cette structure conduit à la notion d'isoenzymes(isozymes) qui sont des formes de la même enzyme catalysant la même réaction, mais dont certaines propriétés physico-chimiques (charge électrique, taille, ..) sont différentes.
Les isoenzymes sont en général mises en évidence par la technique d'électrophorèse. La lactate déshydrogénase (LDH) est l'un des exemples les mieux connus.
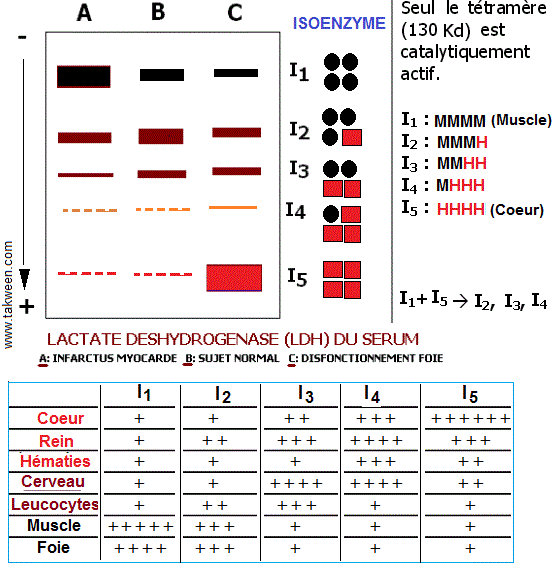
Les isoenzymes sont exploitées dans le domaine d'étiquetage des ressources génétiques animales, végétales et microbiennes (diversité génétique). Elles sont utilisées en génétique des populations vu leur caractère de codominance (les hybrides sont facilement différenciés par rapport aux parents).
QUIZ
Question 1.
Laquelle des substitutions d’acides aminés suivantes serait la moins susceptible d’affecter l’activité d'uneenzyme ?
(!Valine par Aspartate)(!Glutamate par Asparagine)(!Tryptophane par Glycine)(Arginine par Lysine)
Question 2.
About the active site of an enzyme, which of the following statements is correct?
(!The active site binds the substrate more tightly than it does the transition state intermediate)(!The active site binds the product of the reaction more tightly than it does the transition state intermediate.)(The active site binds the substrate less tightly than it does the transition state intermediate.)
Question 3.
Which of the following statements is FALSE regarding the active site of an enzyme?
(An active site is normally hydrophilic. )(! An active site is a cleft on the surface of an enzyme.)(!Substrates fit into active sites and bind to functional groups within the active site.)
Question 4.
Une enzyme dont la séquence d’acides aminés est modifiée connait les changements:
(Modifications possibles au niveau du site actif par altération de la fonction catalytique en permettant toujours la fixation du substrat.)(! Modifications certaine de la structure spatiale et altération de la fonction catalytique en bloquant la fixation du substrat)(!Altération certaine de la catalyse enzymatique.)
Détails des réponses:
Q1. On doit prendre en considération les propriétés des acides aminés contituant l'enzyme ou son site actif. Ainsi, au pH physiologique (pH 7,4), certaines chaînes latérales d’acides aminés portent une charge négative, certaines portent une charge positive et d’autres seront neutres. Le remplacement d'un acide aminé par un autre ne doit pas impliquer des caractéristiques différentes.
- Le remplacement de la Valine par l'Aspartate peut affecter l'activité de l'enzyme. En effet, l'aspartate est chargé négativement au pH physiologique, tandis que la valine est proche du neutre.
- Le remplacement du Glutamate par l'Asparagine peut affecter l'activité de l'enzyme, étant donné que le glutamate (pI=3,2) est chargé négativement, alors que l’asparagine est proche du neutre (pI=5,4), cette substitution interférerait probablement avec l’activité enzymatique.
- La substitution d'un résidu tryptophane par la glycine pourrait potentiellement conduire à des effets stériques susceptibles d'interférer avec la liaison du substrat à l'enzyme. En effet, contrairement à la chaîne latérale extrêmement petite de la glycine, celle du tryptophane est très grande.
- la substitution de l'Arginine (chargée positivement) par une lysine (chargée positivement) serait la moins susceptible de provoquer une perturbation de l'activité de l'enzyme, en raison de la similitude entre ces deux acides aminés.
Q2. For an enzyme reaction, the substrate molecule(s) form a transition state intermediate (when the molecule is neither a substrate or product), which rapidly converts to products because it is unstable. The active site of the enzyme is complementary to the transition state intermediate. This lowers the energy of activation for the reaction thus increasing the reaction rate.
Q3.An active site is normally hydrophobic, in nature.
 |
 |
 |
 |
Vidéo 'Enzymes, catalyseurs biologiques (Ar) الإنزيمات، محفزات بيولوجية'
Liens utiles:
- الإنزيمات. البنيات الوظيفية والموقع النشيط - Enzymes. Functional structures and active site
- MECANISMES MOLECULAIRES DE LA CATALYSE BIOLOGIQUE ET IMPORTANCE DU SITE ACTIF DES ENZYMES
- CATALYSE. INTERACTION MOLECULAIRE ENZYME-SUBSTRAT.
- Formation de complexe enzyme-substrat (ES)
- Cinetique enzymatique et équation de Michaelis-Menten
- Equation de Michaelis-Menten.
- Expressions de l'activité enzymatique
- Inhibition par excès de substrat
- CATALYSE. INTERACTIONS MOLECULAIRES ENZYME SUBSTRAT-
- INHIBITEUR
- INTERACTIONS MOLECULAIRES ENZYME (E)-SUBSTRAT (S)-
- ACTIVATEUR (A)
- ENZYMES ALLOSTERIQUES
- EXAMENS
Ouvrages de la préparation de la transition Secondaire-Supérieur


-
من علوم الحياة إلى البيوكيمياء
Des Sciences de la vie au Lycée à la Biochimie à l'Université
-
تحميل ملفات في مواضيع البيوكيمياء
Téléchargement de fichiers en Biochimie

BIOCHIMIE. TP CONSIGNES (Vidéos)
Chaine
Youtube (abonnement). Plusieurs vidéos multilingues

مجلة إخبارية - Newsletter

ANNONCES, INFORMATIONS
Congrès, Conférences,...
- Congrès, Conférences
- Cours, Workshops
- Bourses, Postdocs
- Biotech-ecolo annonces
CONCOURS:
Concours de
Pharmacie
Afin de pouvoir continuer à servir les visiteurs, soutenez nos actions sur le site takween en faisant acquisition des ouvrages et supports pédagogiques desitinés à améliorer l'enseignement et la recherche scientfique en Biochimie.


Faculty of Sciences, Cadi Ayyad University
Marrakech, 40000, Morocco
Email: baaziz@uca.ac.ma
Phone: 212524434649 (post 513)
Fax: 212524434669
About takween.com